Cytology


Definition and basic characteristics of the cell[edit | edit source]
A cell is the smallest basic structural and functional unit of a living organism.
Most cells are adapted according to structure and function (except undifferentiated cells).
Human cells are 10-30 μm, though some are smaller (erythrocytes: 7.5 μm) or larger (oocytes: 120-150 μm).
The cytoplasmic membrane encloses the cell. It is composed of a phospholipid bilayer with
- embedded proteins
- cholesterol
- oligosaccharides bound to proteins or phospholipids
The basic compartments within the eukaryotic cell are the cytoplasm and the nucleus.
General structure of the cytoplasm[edit | edit source]
The cytoplasm includes all the material within the cell membrane, but excluding the nucleus. The main components include
- cytosol (gel-like fluid where many cellular processes take place)
- organelles
- inclusions
Cell surfaces and intercellular junctions[edit | edit source]
Epithelial cells have apicobasal polarity - organelles and membrane proteins are unevenly distributed within the cell.
Basal pole[edit | edit source]
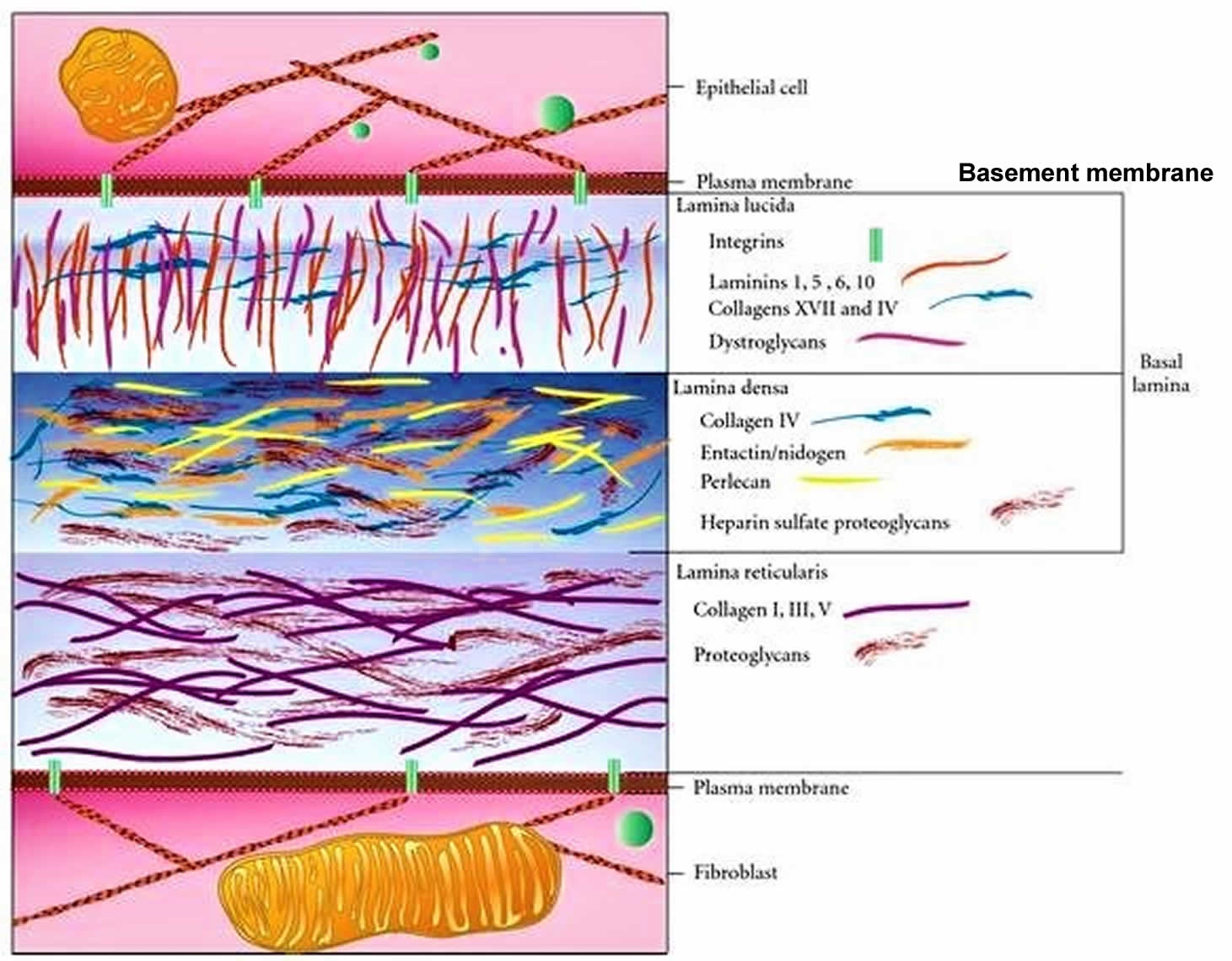
Basement membrane[edit | edit source]
The basement membrane is the means of attachment of the basal poles of epithelial cells with the ECM and surrounding connective tissue. It functions as a selective filter and in communication.
The basement membrane consists of a basal lamina (20-100 nm sheet of dense fibrils) and a reticular lamina (50-500 nm layer of diffuse fibers).
| Layer | Components | |
|---|---|---|
| Basal lamina | lamina rara externa | laminin, perlecan, fibronectin, syndecans, integrins |
| lamina densa | heparansulfate, collagen type IV | |
| lamina rara interna | hyaluronic acid, thrombospondin, fibronectin, collagen type VII | |
| Reticular lamina | collagen types I, III, VII; fibronectin and versican | |
Schematic here.
In some organs, the basal laminae of epithelial layers can fuse. This happens in the Bowman's capsule, cornea, and retina.
Connections[edit | edit source]
Hemidesmosomes (associated with intermediate filaments) anchor the cytoskeleton of the epithelial cells to the basal lamina via intermediate filaments.
Focal adhesions are similar to hemidesmosomes, but are smaller and use integrins instead of intermediate filaments.
If the cells detach, progammed cell death - anoikis - happens.
Basal labyrinth[edit | edit source]
Some epithelia, such as renal proximal and distal tubules, collecting ducts, and striated ducts of salivary glands, also have a basal labyrinth. It consists of infoldings of the basal plasma membrane and interdigitations of basal ridges with neighboring cells. This greatly increases surface area, which is necessary for the transcellular transport that these epitheliae do. Mitochondria accumulate in this area and align vertically[1].
Apical pole[edit | edit source]
The apical pole is opposite to the basal pole and usually has a free surface facing an open space. Specializations include
- microvilli (0.1 μm by 1 μm): projections of cytoplasm supported by microfilaments. Increase surface area to facilitate absorption. Organize together into striated border (enterocytes) or brush border (proximal tubule of kidney).
- stereocilia: similar to microvilli, but longer (10-50 μm). Function in absorption (ex. epithelia of male reproductive tract) and also in sound detection (cochlea).
- kinocilia (0.2 by 5-10 μm): contain axonemal (9×2+2) arrangements of microtubules moved by kinesin and dynein. Found in cuboidal or columnar cells of many epithelia (ex. oviduct); used to move fluid and suspended particles, also function in motion detection in inner ear
- primary cilium: non-motile projection for signal transduction ("antenna") found in almost every vertebrate cell
Lateral surfaces[edit | edit source]
The lateral surfaces touch neighboring cells. There are several types of intercellular adhesions and junctions:
- Tight junctions (aka zonula occludens): tight seal between adjacent cells associated with actin. Most apical.
- Adhesions
- zonula adherens: points for adjacent cytoskeletons to connect and re-inforce tight junctions
- macula adherens (aka desmosome) ~0.3 μm × 20-40 nm: associated with intermediate filaments (keratins). Strong intermediate coupling that strengthens tissue.
- Communicating junctions (aka gap junctions or nexons) ~0.3 μm diameter, pore diameter is <1.5nm, intercellular space is between 2-4 nm: made up of connexins; allow transfer of small molecules between cells
Cytoskeleton[edit | edit source]
Three main kinds of protein polymers make up the cytoskeleton:
- Microfilaments (7 nm):
- G-actin monomers make up F-actin
- Anchored to cell membrane
- Fast polymerization/depolymerization
- Involved in moving the cell, changing its shape, cytokinesis, cytoplasmic transport and streaming.
- Intermediate filaments (10 nm):
- Antiparallel tetramers made of various subunits
- found throughout cytoplasm and in desmosomes
- Highly stable structure; disassembly and reassembly not possible
- Strengthens structure
- Microtubules (25 nm):
- arrangements of 13 heterodimers of α and β tubulin form a hollow tube
- grow from centrosomes
- fast polymerization and depolymerization
- Maintains structure, provides "tracks" for intracellular transport, moves cilia and flagella
Nucleus[edit | edit source]
The nucleus (5-10 μm) stores the cell’s genetic material.
The nuclear envelope forms a boundary continuous with the endoplasmic reticulum. It is made up of two lipid membranes separated by a perinuclear space (30-50 nm) and perforated by nuclear pore complexes (100-125 nm). The lamina (80-100 nm) is a meshwork of proteins (ex. lamins) that strengthens the envelope and serves as anchorage for chromatin. Cryofracture is particularly useful in visualizing the lipid membrane and associated structures.
Chromatin includes the DNA and associated proteins. Euchromatin is transcriptionally active – the chromosomes are uncoiled, and therefore it is invisible with light microscopy. Heterochromatin is transcriptionally inactive, the DNA is coiled around histones and visible as dark dense granules.
The nucleolus is a non-membrane-bound structure, where rRNA is synthesized (pars fibrosa) and ribosomes are assembled (pars granulosa). It disappears during prophase, since the DNA needs to condense into chromosomes.
Organelles[edit | edit source]
Organelles are metabolically active structures within the cytoplasm.
Endoplasmic reticulum[edit | edit source]
- Membranous network of tubuli and vesicles that empty into cisterns
- Continuous with nuclear envelope
- Site of protein synthesis and modification via ribosomes (Rough ER)
- Site of lipid metabolism, glycogen breakdown, drug detoxification (Smooth ER)
Golgi apparatus[edit | edit source]
- Consists of stacks of disk-shaped membranous cisternae
- receives incoming vesicles from the RER at the cis face
- modifies and packages the proteins
- sends proteins in vesicles to their destination from the trans face
Mitochondria[edit | edit source]
- 0.5-1 μm diameter, up to 10 μm long
- Outer membrane encloses intermembrane space
- Inner membrane lies beneath, many folds (cristae) increase surface area and project inward
- Contains own circular mtDNA in matrix
- Functions in the Krebs cycle, oxidative respiration, and apoptotic pathway
Lysosomes[edit | edit source]
- 0.05-0.5 μm
- Membrane-limited vesicles with over 50 types of acidic (pH 5) hydrolases
- Abundant in phagocytic cells
- Degrades excess or nonfunctional components of cytoplasm via autophagy: a lipid bilayer forms around the component and then fuses with a lysosome to form an autophagosome (aka secondary lysosome)
- Contents are recycled in the cytoplasm; the “trash” remains in the residual body as lipofuscin
Peroxisome[edit | edit source]
- Single-membrane vesicle
- contains oxidases that form H2O2 while oxidizing substrates
- catalases break down peroxide
- functions in detoxification and beta-oxidation of fatty acids
Cell inclusions[edit | edit source]
Cytoplasmic inclusions contain substances but are not metabolically active. Most are not enclosed by a membrane. There are several different types:
- Secretory granules: contain compounds destined for secretion out of cell
- Storage compounds
- Glycogen
- glucose polymer linked by α(1→4) glycosidic bonds
- alpha-glycogen granules (up to 200 nm): rosette-like aggregations in hepatocytes
- beta-glycogen granules (15-20 nm): spherical aggregations in skeletal and cardiac muscle[2]
- Lipids
- one large droplet in white adipocytes
- multiple smaller droplets in brown adipocytes
- also present in cells of adrenal cortex and liver
- Protein crystals
- Glycogen
- Pigments
- Endogenous
- autogenous (ex: melanin)
- hematogenous (ex: hemoglobin decomposition)
- Exogenous (from environment; ex: carotenes, inhaled particles)
- Endogenous
- Exocytic vesicles: contain substances targeted for exocytosis; vesicle fuses with membrane and buds off of cell
- Endocytic vesicles: incoming substances get engulfed by membrane that fuse with the membrane; the formed vesicle buds off
Vital manifestation of the cell[edit | edit source]
- Movement (of the cell itself, of substances outside and inside it)
- Metabolism
- Response to stimuli
- Growth
- Differentiation
- Division
References[edit | edit source]
Mescher, A. and Junqueira, L., 2018. Junqueira's basic histology. New York: McGraw-Hill, pp. 17-68

{kind=link}